So, here is the premiere of Darwin Eats Cake:

URL for embedding: http://www.darwineatscake.com/img/comic/1.jpg
For the latest, go to the website.
So, here is the premiere of Darwin Eats Cake:
URL for embedding: http://www.darwineatscake.com/img/comic/1.jpg
For the latest, go to the website.
So, it’s Sunday morning here in the States. If you’re reading this now, it means you’re skipping church, which means you might also enjoy this awesome video by Ben Hillman. I came across it via Boing Boing.
Evolution Made Us All from Ben Hillman on Vimeo.
So, our next scientific Egypt Week post concerns a paper just published in last week’s issue of Nature, where the authors describe novel behavioral effects of the imprinted gene Grb10 in the mouse.
If you’re not familiar, genomic imprinting is the phenomenon where the expression pattern of a gene depends on its parental origin. So, most of your genes come in two copies, one of which came from your mom, and one of which came from your dad. For most genes, the function of the allele, or gene copy, depends just on its DNA sequence. But something like 1% of our genes are imprinted, meaning that they retain a chemical memory of which parent they came from, so that the two gene copies will function differently, even if the DNA sequences are identical.
The most widely accepted theory for the evolutionary origin of gene expression suggests that it is the result of an intragenomic conflict between maternally and paternally inherited gene copies. That is, from a gene’s-eye point of view, natural selection acts differently on maternally and paternally derived alleles.
Many imprinted genes in mammals have growth effects in early development, and these most of these effects are well described by models where selection favors more growth (and a greater demand on maternal resources) when alleles are paternally derived, and less growth (preserving more maternal resources for the mother’s other offspring) when maternally derived.
There is also evidence for large-scale imprinted gene expression in the brain, and evidence that these imprinted genes may have substantial effects on cognition and behavior. We are still at the early stages of describing these effects, and at even earlier stages of understanding the relevant evolutionary pressures.
Elsewhere on this blog, I have begun writing a series of primers on genomic imprinting, links to which can be found here, if you are interested in more background.
Today’s paper describes the effects of the two parental knockouts of the Grb10 gene. Grb10 is a particularly interesting imprinted gene, because it is maternally expressed in many peripheral tissues, but paternally expressed in the central nervous system. So, when you knock out the maternally inherited copy, you get a complete loss of function in the periphery, but don’t impact Grb10 expression in the brain. Conversely, when you knock out the paternally inherited copy, you lose gene function in the brain, but leave expression in the periphery unaffected.
The phenotype of the maternal knockout is more or less what is expected in terms of growth effects, and is consistent with previous studies of this gene. Theory predicts that if a growth-related imprinted gene is maternally expressed, it likely functions as a suppressor of growth. When the maternal copy of Grb10 was knocked out, the result was overgrowth, due to the loss of this growth-suppressing function.
The knockout of the paternally inherited results in a behavioral phenotype associated with increased social dominance, as indicated by two specific behaviors. The first dominance behavior was observed in a “tube test.” In this test, two mice who don’t know each other are forced to encounter each other in a tube. In this setting, the knockout mice are less likely to back down than the wild-type (normal) mice are.
The second observation was an increase in allogrooming and barbering. Let’s pause for a moment to talk about what that means. Allogrooming is where one individual grooms another individual (in contrast to autogrooming, where you groom yourself). Barbering is where the grooming gets out of hand, and the groomee gets big bald (and sometimes bruised and bloody) patches.
Now, intuitively, you might assume that grooming behavior is submissive, like the handmaid combing out the princess’s hair. In mice, at least, it’s not like that. If you have a pet mouse, and it is grooming you, it is actually being dominant. It’s more like when you sit your little sister down in a chair and put makeup on her – the goal is NOT to make her look good. And, if you are feeling really mean, you give her a haircut, too.
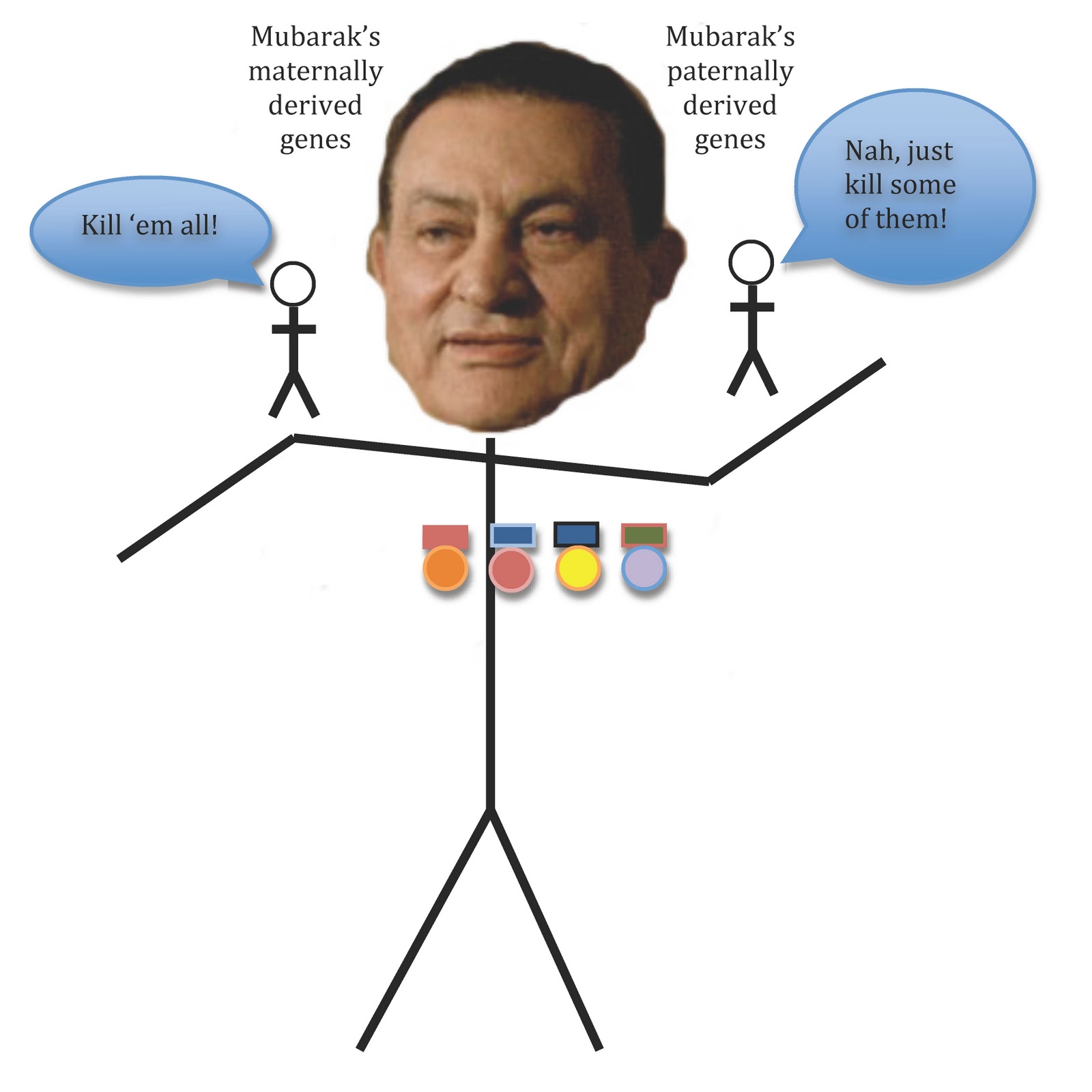 |
| A bemedallioned Hosni Mubarak helps to illustrate the intragenomic conflict over social dominance behaviors. Natural selection favors alleles that enhance socially dominant behaviors when they are maternally derived, but limit socially dominant behaviors when paternally derived. The study was performed in mice, and it is important to note that the patterns of imprinted gene expression can vary among species, so we can not extrapolate from these results to the influence of Grb10 on human cognition and behavior. However, mice and rats are closely related, so we are probably safe extrapolating to Mubarak. |
The next question is why would alleles favor more socially dominant behaviors when maternally derived? Fundamentally, at this point we have no idea. This is where the modeling has to come in. In this type of situation it is always possible to come up with a host of possible explanations, all of which sound plausible, and all of which would predict that a paternally expressed gene would limit dominance. The key thing is to model each of those explanations formally, so that we know what key ecological and demographic factors underlie the explanation. Then, we find other species where those factors differ, and examine the imprinting status and phenotypic effect of Grb10 in those species.
 |
| For the less politically oriented, the intragenomic conflict over social dominance is like this. Nadya “Octomom” Suleman is like your maternally inherited genome, while the guy with the moustache and the milk bottle is like your paternally inherited genome. Image from the Daily Mail. |
Peace be upon you.
Garfield AS, Cowley M, Smith FM, Moorwood K, Stewart-Cox JE, Gilroy K, Baker S, Xia J, Dalley JW, Hurst LD, Wilkinson LS, Isles AR, & Ward A (2011). Distinct physiological and behavioural functions for parental alleles of imprinted Grb10. Nature, 469 (7331), 534-8 PMID: 21270893 [1]
––––––––––––––––––––––––––––––––––––––––––––––––––––––
[1] Disclosure: I didn’t really intend for Egypt Week to devolve into blog-posts-about-papers-by-collaborators-of-mine week, but there you have it. I have an ongoing collaboration with Anthony Isles, and know some of the other authors.
![]() So, our next Egypt Week feature is a theoretical paper on a topic closely related to the last post. Once again, we are interested in understanding the mechanisms that are responsible for encouraging or enforcing cooperation, thereby facilitating collective action. Last time, we talked about a paper that found that “altruistic” or “third-party” punishment is common in large-scale, complex societies, but is rare in small-scale societies, while “spiteful” punishment is universal.
So, our next Egypt Week feature is a theoretical paper on a topic closely related to the last post. Once again, we are interested in understanding the mechanisms that are responsible for encouraging or enforcing cooperation, thereby facilitating collective action. Last time, we talked about a paper that found that “altruistic” or “third-party” punishment is common in large-scale, complex societies, but is rare in small-scale societies, while “spiteful” punishment is universal.
Many empirical and theoretical studies of cooperation focus on punishment as a mechanism for enforcing societal norms. Basically, you set up a situation where the group benefits if people cooperate, but each individual benefits by not cooperating. If mechanisms exist to punish people for not cooperating, you get cooperation. Which is to say that the existence of punishment changes the individuals’ incentives. The benefits of not cooperating are outweighed by the cost of being punished. No big mystery there.
But what if punishment itself is costly? Punishment can stabilize cooperation, but what stabilizes punishment? Some models rely on an infinite succession of punishments, where people punish people who fail to punish people who fail to cooperate, and people punish people who fail to punish people who fail to punish people who fail to cooperate, and people punish … well, you get the idea.
Today’s paper asks if cooperation can be enforced by corrupt punishment. That is, while punishment is still treated as costly, punishers are not necessarily cooperators themselves, as is commonly assumed in models of this sort. Furthermore, the corrupt punishers (“policers”) suffer a lower cost when punished than do non-punishers (“civilians”).
 |
| A corrupt policer looks forward to a cushy retirement, thanks to his hypocritical enforcement of others’ cooperation. Little does he suspect how a new, young partner, who colors outside the lines, but has a heart of gold, will turn his whole life upside-down, with hilarious consequences. |
The model shows that in the presence of a modest power imbalance, cooperating civilians and corrupt policers can coexist. That is, a moderate level of corruption is consistent with, and can even stabilize cooperation. However, when the power imbalance becomes large, corrupt policers overrun the population, the system breaks down, and cooperation is lost.
The first part of the result is nice because it provides a degree of robustness to the “cooperation through punishment” paradigm, as it does not require the punishers to be acting altruistically themselves.
The second part of the result is perhaps more directly relevant to Egypt Week. Societies can function in the presence of a degree of inequality, and they can tolerate a certain amount of hypocrisy from their leaders. But too much hypocrisy and inequality is inconsistent with the type of collective action that governments are meant to facilitate.
It is heartening to see that when a less corrupt alternative presents itself, people are still capable of collective action on a massive scale.
Peace be upon you.
Úbeda, F., & Duéñez-Guzmán, E. (2010). POWER AND CORRUPTION Evolution DOI: 10.1111/j.1558-5646.2010.01194.x [1] [2]
––––––––––––––––––––––––––––––––––––––––––––––––––––––
[1] This is an online, ahead-of-print publication, which is why there are no page numbers, but it should be findable through the DOI.
[2] Disclosure: The first author on this paper is a long-time friend and colleague, and we have worked together on issues of intragenomic conflict. Here is photographic evidence of our friendship, from when we were traveling around Lyon, France like Thelmo and Louis following the 2010 SMBE meeting:
 |
| On our way to the Palais de Justice, we accidentally activated our Wonder-Twin Powers. Francisco took the shape of an evolutionary biologist, and I took the form of a French trash can. Photo by Gleek. |
So, welcome to the first Egypt Week edition of Lost in Transcription. We’re going to kick it off with an anthropology paper that uses a cross-cultural approach to study the origins of human punishment and cooperation.
If you’re not familiar with this vein of research, let me set the stage for you. The “problem” of cooperation when people talk about it in anthropology, biology, and economics is this. If you take a super naive view of natural selection, it would say that we should have evolved to ruthlessly pursue our own self interest. In particular, if we have an opportunity to cheat and get away with it, the logic of self interest suggests that we should. From this perspective, the whole idea of successfully engaging in collective action seems absurd.
Contrary to this naive expectation, we observe that people do forego opportunities to pursue their own narrow self interest, and the history of civilization is one of successful collective action on an enormous scale.
Of course, at some level, we know what the answer is.[1] Natural selection does not act only on the short-term self interest of the individual, but favors behaviors that enhance survival, reproduction, and the propagation of the genes carried by the individual. Those things are all affected by more that just who gets the biggest piece of the pie in a given interaction. Other factors that come in to play include selection on kin or social groups, the establishment and enforcement of social norms, and systems of reward, punishment, and reputation.
So, much of the work in this field focuses on trying to figure out which among these various effects has played the greatest role in the origin of the enormous capacity for cooperation that underlies all human societies.
This paper uses a standard set of experimental protocols, applied to twelve societies that differ enormously in size and complexity of social organization. The societies studied span the range from the Hadza, hunter-gatherers from Tanzania, to the people of Accra, the capital city of Ghana. Each experiment is a two-player or three-player game in which the players make decisions that determine how money is distributed. The goal here is to measure the degree of “altruism” in each society, the degree of “second-party” punishment (how willing are people to punish someone who is not generous to them), and the degree of “third-party” punishment (how willing are people to punish someone who is not generous to someone else).
The first experiment used was the Dictator Game (DG), in which the experimenter provides Player 1 with an allotment of money, and Player 1 determines how that money will be divided between themselves and Player 2. The game is played once, and is played anonymously, so there is nothing to stop Player 1 from offering nothing to Player 2. This experiment establishes a baseline level of cooperation or altruism, quantified by the average proportion of the money that Player 2 receives.
The second experiment is the Ultimatum Game (UG). This is like the Dictator Game, but Player 1 proposes a division of the allotment of money, and Player 2 can either accept or reject this proposal. If Player 2 rejects, both players walk away with nothing. This measures the willingness to punish ungenerous individuals who make low offers. Note that for any non-zero offer, Player 2 actually has to give up money in order to punish Player 1, making this a “spiteful” form of punishment.
The third experiment is the Dictator Game, but involves a third player, who receives an allotment of money from the experimenter. After Player 1 determines how the primary allotment will be divided between him/herself and Player 2, this third player has the option of paying back a portion of their allotment in order to have three times that amount taken away from Player 1. This again measures the willingness to punish an ungenerous offer, but this time the punishment is “altruistic” rather than “spiteful,” since the punisher was not actually the one who suffered the ungenerous offer.
A few general patterns emerged from this set of experiments.
(1) Small-scale societies were less generous (Player 1 made lower offers) across all three games compared with larger societies.
(2) Second-party punishment was observed at similar rates across all societies. Thus, in the small-scale societies, individuals “expect to get a fair share even when they do not want to give a fair share.”
(3) Third-party punishment was much more common in larger societies. And, in fact, in those larger societies, third-party, “altruistic” punishment occurred more often than “spiteful” second-party punishment.
Taken together, what these patterns support the idea that human cooperation may have emerged first in the context of spiteful punishment, rather than through altruistic or community-oriented enforcement of social norms. They suggest that third-party punishment arises only with the establishment of more complex societies. In particular, once a society exceeds a certain size, it becomes difficult to keep track of individual reputations. In such groups, collective-action problems require the existence of institutions that promote and reward third-party punishment.
Peace be upon you.
Marlowe, F., Berbesque, J., Barrett, C., Bolyanatz, A., Gurven, M., & Tracer, D. (2010). The ‘spiteful’ origins of human cooperation Proceedings of the Royal Society B: Biological Sciences DOI: 10.1098/rspb.2010.2342 [2]
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
[1] Editorial note: This is a version of the answer that comes up again and again in science. The world is a complicated place. We (scientists) come in and try to describe it using a really simple model. Then we feign surprise that the simple model does not adequately describe the behavior of the system that we are supposedly modeling. We declare that there is a paradox and build a field around resolving it. This is all fine, as it is just the way that science works, but it is worth remembering that many of the “problems” in science are really artifacts of simplistic models that have achieved an iconic status.
[2] Disclosure: Frank Marlowe, who is the first author on this paper, is a friend, and we have written a paper together on the genetic and ethnographic evidence regarding sex-biased migration in humans.
So, unless you are in China, you are undoubtedly aware of the massive protests that have been going on in Egypt for the past few days. If you’re like me, you are filled with a mixture of hope and anxiety. I sincerely hope that the Egyptian people will be able to establish a new government that is actually responsive to their needs, and that it can be done with a minimum of bloodshed. But I also know that it is possible that this ends with a brutal crackdown and the Egyptian government resuming business as usual.
Also, if you’re like me, you feel a sense of solidarity with the protesters, but feel completely powerless to do or contribute anything. I am thousands of miles away, and do not have any inside information or clever insights on Egyptian politics and culture. I am a poet and a theoretical evolutionary biologist – two things that find little practical application even in the best of circumstances.
Still, I feel that I want to do something, so I will do what I know. I am going to dedicate this week’s blog entries to the protesters in Egypt and to people everywhere who long for more freedom and better government. Topic-wise, I will discuss a number of recent scientific papers from the evolutionary literature on cooperation, punishment, and corruption.
I have no illusion that this can have any impact on the course of events in Egypt, or on the protests in Jordan, Yemen, and elsewhere. I simply hope that I can contribute something to the knowledge that we are all in this together.
The desire for freedom and safety and a better life for ourselves and our children is not defined or limited by nationality or religion or race or language. As an American, I am fully aware that my government’s rhetoric about freedom and democracy is often at odds with its support of corrupt and anti-democratic regimes, including Mubarak’s. But a government is rarely the same thing as a people. All of the people I have spoken with, American and not, are hoping and praying that this will be a turning point in history, and that the people of Egypt (and Iran and Myanmar and Uganda and on and on) can move forward together into a better future.
Peace be upon you.
So, we’ve already discussed the fact that genomic imprinting is mediated through epigenetic differences between the maternally and paternally inherited gene copies. That is, at an imprinted locus, the maternally inherited allele will have one pattern of epigenetic modifications, while the paternally inherited allele has a different pattern. These differences are first established in the male and female germ lines, when the alleles that will eventually become maternally and paternally derived are in physically different locations. It is not hard to imagine, then, how these differences could be established. One pattern of gene expression in spermatogenesis results in the paternal-specific epigenetic modifications. A different pattern of gene expression in oogenesis results in maternal-specific epigenetic modifications.
But what are these epigenetic modifications, and how do they change the expression pattern of the gene?
There are a number of modifications involved in imprinting, but for the moment, we’re going to focus specifically on the simplest and best-understood mechanism: DNA methylation.

The two horizontal lines in this picture represent the two copies of a gene. The big, solid box is the part of the gene that actually codes for the protein. The open box is the promoter region, which is the part of the DNA sequence responsible for regulating expression of the gene. The lollipop things indicate DNA methylation on cytosine residues (the “C” of the A, C, G, T alphabet that makes up DNA).
In this simplest type of scenario, the DNA sequence in the promoter region binds to a variety of proteins that recruit the molecular machinery that will transcribe the gene, leading eventually to production of the corresponding protein. The addition of methyl groups to the DNA changes its binding properties, so that it no longer binds to this machinery, and that copy of the gene is not transcribed.
If you’re not a molecular biologist, you can think of it like this. The transcription machinery is a bit like a Xerox machine, and the gene is like the master copy of some document. The promoter region is like a lock that has to be unlocked before you can copy this particular document. There are a number of proteins called “transcription factors” that function like a key to this lock. These transcription factors fit nicely on the promoter region, unlocking the gene and resulting in the production of many copies of the gene product.
Adding methylation to the promoter region is a bit like squirting epoxy into the lock. The presence of the methyl groups actually changes the physical shape and chemical properties of the DNA. So, when you try to put the key in, it no longer fits right, and the gene can not be copied.

In the top part, we see the red transcription factor binding to the black promoter region, which will activate transcription from the gene. In the bottom part, methyl (CH3) groups have been chemically added to the promoter region, preventing binding, and thereby preventing transcription.
So, these relatively subtle chemical changes are able to completely alter the functional properties of the gene.
Next time, we’ll talk about how these methylation patterns are maintained through development, and how the two gene copies are able to maintain distinct epigenetic states across multiple rounds of cell division and DNA replication. Make sure to tune in, because it’s really slick!
The two references represent the first proposals that DNA methylation might be the thing that permits the stable transmission of patterns of gene expression across cell divisions.
Holliday, R., & Pugh, J. (1975). DNA modification mechanisms and gene activity during development Science, 187 (4173), 226-232 DOI: 10.1126/science.1111098
Riggs, A. (1975). X inactivation, differentiation, and DNA methylation Cytogenetic and Genome Research, 14 (1), 9-25 DOI: 10.1159/000130315
So, in the previous installment, we introduced the “Loudest Voice Prevails” principle, which describes the evolutionarily stable pattern of gene expression at an imprinted locus where there is an intragenomic conflict over the total level of gene expression. Basically, the allele that favors lower expression becomes transcriptionally silenced. Expression from the other allele (the “louder” voice) evolves to the level that maximizes its inclusive fitness. In this sense, the active allele at an imprinted locus “wins.”
But what is going to happen if we have a pair of imprinted genes that exert opposite effects on the phenotype? If we have a paternally expressed growth enhancer, it will evolve to bring the growth phenotype up to the paternal optimum. If we have a maternally expressed growth suppressor, it will evolve to bring the growth phenotype down to the maternal optimum. But what if we have both?
Well, intuitively, if there is conflict between maternally and paternally derived genes over the optimal growth phenotype, then the phenotype can’t simultaneously satisfy the paternal and maternal optima. One or the other (or both) of these genes will always be under selection to increase its gene expression level (or, equivalently, the activity or longevity of the gene product, etc.). Thus, these two opposing genes will become involved in a kind of arms race.
In the simplest possible model that we can write down, this arms race goes on indefinitely, with natural selection driving each of the genes towards infinite expression. Clearly, in a real biological situation, this will not be the case, and something will step in to bring this escalation to a halt. The questions then become: What stops the escalation? And, what does the system look like at its new, escalated, evolutionarily stable state?
To think about this, let’s return to our analogy from last time, where Pat and Chris are sharing an office, but disagree about what temperature the office should be kept at. Recall that genes are totally passive aggressive, so Pat and Chris don’t compromise or communicate. They just use the tools at their disposal to move the office closer to their preferred temperature. Pat wants the office at 71 degrees. Chris wants it at 70.
We saw that if Pat and Chris both have space heaters, eventually Chris’s space heater is off, while Pat’s holds the temperature at 71. On the other hand, if they both have air conditioners, Pat will turn his/her A/C off, and Chris will get to have the room at 70.
If each of them has a space heater and an air conditioner, we have an arms race on our hands. Whenever the temperature is below 71, Pat will turn up the space heater. Whenever it is above 70, Chris will turn up the air conditioner. In passive-aggressive-allele fashion, this will go back and forth until the space heater and air conditioner are both blasting away. In the absence of any constraints or side effects, it will go on until both are blasting away infinitely.
There are several ways that the escalation could stop, however, each of which has a biological analog.
(1) Mechanical limitation. There will be some limit beyond which gene expression / activity can not increase. Once one of the genes reaches its limit, the other will win. Like if Pat’s Tufnel-brand space heater goes to eleven, Pat wins. Of course, this will depend on the mechanisms through which the two genes exert their influence. For instance, if Chris’s air conditioner is actually a combination air conditioner / food processor / exfoliator, Chris might have to turn it way way up to get the air conditioning equivalent of a little bit of space heating. Similarly, a gene product might perform multiple tasks, and this pleiotropy could limit its competitive ability in the arms race.
(2) Production costs. One difference between the single-locus solution and the two-locus solution is the level of energy consumption. If Chris’s space heater is off, Pat’s holds the temperature at 71. If Chris’s air conditioner is maxed out at ten, Pat’s space heater (which goes to eleven, remember) holds the temperature at 71. The difference is that the second solution comes with a huge-ass electricity bill. Can this sort of cost actually halt the escalation? Maybe. This requires either that there are diminishing returns to increased escalation, or that there are accelerating costs to production (like utility rates where your thousandth kilowatt-hour costs more than your first one).
(3) Intervention. In a real office, we might expect that the manager would come in and yell at Pat and Chris, telling them to turn down their space heater and air conditioner. Maybe the manage would mandate an office temperature of 70.5 degrees. Does this ever happen with genes? Could a consortium of unimprinted genes step in and stop the escalation? There is no evidence to my knowledge of such things happening in the context of genomic imprinting, but this type of intervention is thought to be responsible for meiotic sex-chromosome inactivation, where the autosomes all gang up and put the sex chromosomes in a headlock in order to prevent meiotic drive.
(4) Side effects. What if turning up Pat’s space heater also makes the music louder in the office? What if Chris’s air conditioner draws so much power that it causes occasional brown-outs? This is the other way in which the escalation between imprinted genes might be self limiting. If we consider a monolithic “growth phenotype” in isolation, then each allele has a simple, monolithic optimum. But genes are seldom like that. A paternally expressed allele may benefit from increased expression due to the effect of that increased expression on growth. But what if that increased expression has other consequences, as well? Maybe those other effects are detrimental to the allele’s inclusive fitness. If those deleterious side effects outweigh the growth-related benefits, then natural selection will not drive further escalation.
In future installments, we’ll look at some specific examples of escalating genes. But first, we’ll step back and look at some of the other features and consequences of imprinted genes.
Kondoh, M., & Higashi, M. (2000). Reproductive Isolation Mechanism Resulting from Resolution of Intragenomic Conflict The American Naturalist, 156 (5), 511-518 DOI: 10.1086/303409
Wilkins, J., & Haig, D. (2001). Genomic imprinting of two antagonistic loci Proceedings of the Royal Society B: Biological Sciences, 268 (1479), 1861-1867 DOI: 10.1098/rspb.2001.1651
So, it’s been a while since the last installment of the Primers on Imprinting feature, but they should be posted with greater regularity in the upcoming weeks. This time we’re going to introduce something that we will see again in future installments: small differences in selection lead to large differences in behavior.
Last time, we introduced the most widely discussed and most successful explanation of the evolutionary origins of genomic imprinting, the “kinship” or “conflict” theory. According to this theory, imprinted gene expression is a consequence of the fact that natural selection acts differently on alleles depending on their parent of origin. There are several ways to think about the origin of this differential selection, but we talked about it in terms of the framework that I find most intuitive: inclusive fitness.
As we also noted last time, even in the cases where the asymmetry in selection on maternally and paternally derived alleles is sufficiently large to drive the evolution of imprinted gene expression, the actual magnitude of this asymmetry is actually incredibly small. Why? Well, even for a allele with large effects on the survival and reproduction of related individuals, the dominant factor in the inclusive fitness of that allele is still going to be the survival and reproduction of the individual organism carrying that allele around.
But, the standard pattern observed with imprinted genes is that the allele-specific expression is all or nothing. For example, an allele might be expressed when it is inherited from a male, but completely silent when inherited from a female. So this small difference in the optimal expression levels of the maternally and paternally derived alleles leads to – in a way – the largest possible difference in the realized expression levels of the two alleles.
I like to think of this in terms of an analogy. Imagine that Pat and Chris share an office, and that they have a slight disagreement over the temperature they want the office at. Say Pat wants the office to be at 71 degrees (Fahrenheit), while Chris wants it to be 70. Each of them has control over a small space heater, and this is the only mechanism that they have for manipulating the temperature. [1]
What’s going to happen? Let’s say the temperature is 70 degrees. Pat will turn up his/her space heater until the temperature reaches 71. In response, Chris will turn his/her space heater down until the temperature comes down to 70. They will go back and forth like this until, eventually, Chris’s space heater is completely turned off. Pat will then turn his/her space heater up to get the room to 71. Then we’re done. Chris is unhappy about the temperature of the room, but no longer has any ability to make it any cooler.
Two things about this outcome. First, a small disagreement over the ideal temperature has led to a large divergence in the strategies: Chris’s space heater is all the way off, while Pat’s is on and doing all the work. Notice that the outcome would be exactly the same, in principle, if Chris’s ideal temperature were 70.9 degrees, or even 70.999 degrees. [2]
Second, Pat wins. This is a consequence of the fact that we are talking about space heaters, and that Pat prefers the higher temperature. If, instead of space heaters, Pat and Chris each had control of an air conditioner, Chris would be the winner. At equilibrium, Pat’s air conditioner would be all the way off, and the room would be at 70 degrees.
This is also the way it works with alleles at an imprinted locus. Let’s consider the case of a gene where increased expression results in increased prenatal growth. The inclusive fitness argument says that the optimal amount of this growth factor is higher for an allele when it is paternally inherited than when it is maternally inherited. Say this patrilineal optimum is 105 units, while the maternal optimum is 95 units.
If the gene is not imprinted we might expect it to produce about 100 units, with each allele producing 50. However, once the evolutionary dynamics of imprinting take over, the pattern of expression will evolve to one where alleles are transcriptionally silent when maternally inherited, but where a paternally expressed allele is making 105 units.
For a growth-suppressing gene, where increased expression actually reduces prenatal growth, we expect the opposite pattern, where alleles are silenced when paternally inherited, but are expressed when maternally inherited. This set of predictions – that imprinted growth enhancers will be paternally expressed, and imprinted growth suppressors will be maternally expressed – matches the empirically observed pattern by and large, although there are a few counterexamples that are not fully understood at the moment.
This pattern of allele silencing has been dubbed the “loudest voice prevails” principle. The phenotype evolves to the optimum of the allele favoring higher expression. Now, you can argue that this is the sort of thing that does not really need its own name. Fair enough. It’s really just saying that the evolutionarily stable state of the system is an edge solution. But, “loudest voice prevails” is sort of catchy, and has the advantage of reminding us which allele is expressed at equilibrium.
The Haig 1996 citation is the paper that introduces the phrase. The other three citations are papers published around the same time that use different mathematical frameworks to address the evolution of gene expression at an imprinted locus. Generically speaking, the answer is the one described here, although the Spencer, Feldman, and Clark paper identifies certain regimes in parameter space where apparently different results can be obtained. In a future post, we will delve into the differences in the assumptions and conclusions of different modeling frameworks as they have been applied to imprinting.
Now, what if you consider more than one imprinted gene? What if Pat and Chris each have a space heater and an air conditioner? We’ll talk about that next time.
Haig, D. (1996). Placental hormones, genomic imprinting, and maternal-fetal communication Journal of Evolutionary Biology, 9 (3), 357-380 DOI: 10.1046/j.1420-9101.1996.9030357.x
Mochizuki A, Takeda Y, & Iwasa Y (1996). The evolution of genomic imprinting. Genetics, 144 (3), 1283-95 PMID: 8913768
Haig, D. (1997). Parental antagonism, relatedness asymmetries, and genomic imprinting Proceedings of the Royal Society B: Biological Sciences, 264 (1388), 1657-1662 DOI: 10.1098/rspb.1997.0230
Spencer HG, Feldman MW, & Clark AG (1998). Genetic conflicts, multiple paternity and the evolution of genomic imprinting. Genetics, 148 (2), 893-904 PMID: 9504935
–––––––––––––––––––––––––––––––––––––––
[1] Of course, in the real-life situation, we might assume that Pat and Chris would discuss the situation and come to some sort of agreement. This is a key difference between people interacting in strategic situations and genes evolving under natural selection. Alleles at a locus are like people sharing an office, where both of them are incredibly passive aggressive. If it helps, imagine that Pat and Chris won’t talk to each other.
[2] In practice, of course, there is going to be some minimum level of disagreement required in order to trigger this passive-aggressive escalation. In this analogy, the minimum level will be set by a combination of things such as the sensitivity of Pat and Chris to small changes in temperature, the precision with which the space heaters control the temperature of the room, and the extent to which they care about each other’s comfort. Similar reasoning holds in the case of genes, and we will address this in a future installment of the series, where we ask why there are any genes that are not imprinted.
So, if you haven’t already, you’ll probably soon receive the credit card bill with all of your Christmas purchases on it. Was it worth it? Well, was it, punk?
If you’re like most people, some of your presents were probably intended to impress someone. The question is, what’s the best kind of present for that? Should I give the girl from math class diamond earrings, or new batteries for her calculator? Should I give my boss a mug, or a gift certificate to Glamour Shots?
Fortunately, Science!™ has the answer. Today’s journal club entry concerns a model of gift-giving that considers three different types of gift that differ in their cost to the giver and their value to the recipient. “Cheap” gifts are, well, cheap. “Valuable” gifts are expensive to give, and have value to the recipient. The interesting category is the third one, the “extravagant” gifts, which are expensive to give, but have little inherent value to the recipient.
The specific context is gift-giving and mating. The model is of a sequential game with three or four stages. First, the male offers a gift to the female. Second, the female either accepts or rejects the gift. Third, she chooses whether or not to mate with the male. Then, in one version of the game, the male decides whether or not to stick around and contribute to the care of the offspring.
 |
| This $305 luxury frisbee is an example of an extravagant gift. |
The conclusion of the paper is that there are many combinations of parameter values that will lead to males giving extravagant gifts. There are two critical features of the model that seem to be necessary in order to get this result.
First, there is uncertainty. The female has a guess about the quality of the male (or, equivalently, in the version of the model with paternal care, the probability that he will stick around after mating). By accepting the gift, she gains additional information about his quality or intentions. Similarly, the male is uncertain about the quality and intentions of the female – whether it is worth it for him to stick around after mating, and whether or not she is a gold-digger, who will just take his gift and skip town with his cousin.
[Editorial note: the term “gold-digger” is from the paper. Those of you who know me know me know that I would never have gone with such a politically incorrect term. I would have used “■■■■■■■■■■”.]
[[Meta-editorial note: parts of the previous editorial note have been redacted.]]
The other key feature is that there must be some cost to the female in accepting the gift.
Now, there are lots of parameters in a model like this, and several equilibrium solutions are possible. The interesting one is the one where males give cheap gifts to unattractive females (females whom they judge, with some uncertainty, to be of low quality), and give extravagant gifts to attractive females.
The key to getting the interesting equilibrium is that the ability or willingness to provide and extravagant gift has to correlate with the male’s quality or intentions. For example, a male can’t afford to spend two-months salary on a diamond ring every time he wants to have a one-night stand. Therefore, an extravagant engagement ring becomes a reliable indicator of his intentions. Ideally, the gift has to have no inherent value to the female, for example, if it were impossible to sell the engagement ring for cash money. Recall also that it has to cost her something to accept the gift. Then, taking the gift constitutes a commitment on her part as well. Otherwise, she benefits most from accepting the gift and walking away.
In the salacious application-to-human-mating case, this cost to the female is easiest to envision as a reputation cost (e.g., the risk of being labeled as a ■■■■■■■■■■). In certain species, where females mate with multiple males, store the sperm, and then use it selectively, there may be direct opportunity costs that do not require catty moralizing.
Just one last point.
The paper starts with, “Gift-giving is a feature of human courtship”. The authors cite Geoffrey Miller’s 2000 book, The Mating Mind. If the paper were being written today, I assume they would have cited more recent work by Hefner and Harris.
Sozou, P., & Seymour, R. (2005). Costly but worthless gifts facilitate courtship Proceedings of the Royal Society B: Biological Sciences, 272 (1575), 1877-1884 DOI: 10.1098/rspb.2005.3152